Timosina alfa 1 para uso en el tratamiento de la enfermedad de injerto contra huésped.
Timosina alfa 1 para uso en la prevención o tratamiento de la enfermedad de injerto contra huésped en el trasplante de órganos en mamíferos.
Tipo: Patente Internacional (Tratado de Cooperación de Patentes). Resumen de patente/invención. Número de Solicitud: PCT/EP2007/053321.
Solicitante: SIGMA-TAU INDUSTRIE FARMACEUTICHE RIUNITE S.P.A..
Nacionalidad solicitante: Italia.
Dirección: VIALE SHAKESPEARE, 44 00144 ROME ITALIA.
Inventor/es: GARACI, ENRICO, BISTONI,FRANCESCO, ROMANI,LUIGINA.
Fecha de Publicación: .
Clasificación Internacional de Patentes:
- A61K38/22 NECESIDADES CORRIENTES DE LA VIDA. › A61 CIENCIAS MEDICAS O VETERINARIAS; HIGIENE. › A61K PREPARACIONES DE USO MEDICO, DENTAL O PARA EL ASEO (dispositivos o métodos especialmente concebidos para conferir a los productos farmacéuticos una forma física o de administración particular A61J 3/00; aspectos químicos o utilización de substancias químicas para, la desodorización del aire, la desinfección o la esterilización, vendas, apósitos, almohadillas absorbentes o de los artículos para su realización A61L; composiciones a base de jabón C11D). › A61K 38/00 Preparaciones medicinales que contienen péptidos (péptidos que contienen ciclos beta-lactama A61K 31/00; dipéptidos cíclicos que no tienen en su molécula ningún otro enlace peptídico más que los que forman su ciclo, p. ej. piperazina 2,5-dionas, A61K 31/00; péptidos basados en la ergolina A61K 31/48; que contienen compuestos macromoleculares que tienen unidades aminoácido repartidas estadísticamente A61K 31/74; preparaciones medicinales que contienen antígenos o anticuerpos A61K 39/00; preparaciones medicinales caracterizadas por los ingredientes no activos, p. ej. péptidos como soportes de fármacos, A61K 47/00). › Hormonas (derivados de pro-opiomelanocortina, pro-encefalina o pro-dinorfina A61K 38/33, p. ej. corticotropina A61K 38/35).
PDF original: ES-2553267_T3.pdf

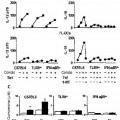

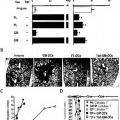
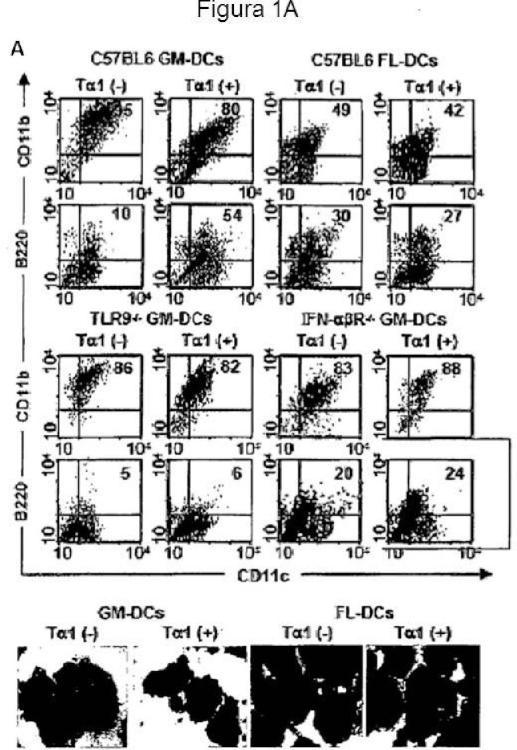
Patentes similares o relacionadas:
Composiciones que comprenden una proteína de fusión de VIP-ELP para su uso en el tratamiento de fibrosis quística, del 29 de Julio de 2020, de Phasebio Pharmaceuticals, Inc: Una composición farmacéutica que comprende una proteína de fusión que comprende un péptido intestinal vasoactivo (VIP) y uno o más péptidos de tipo elastina (ELP), que […]
Antagonistas peptídicos de la familia calcitonina CGRP de hormonas peptídicas y su uso, del 17 de Junio de 2020, de Soares, Christopher, J: Un antagonista del péptido relacionado con el gen de la calcitonina, o una sal farmacéuticamente aceptable del mismo, teniendo dicho antagonista la estructura […]
Derivados de celastrol para el tratamiento de la obesidad, del 3 de Junio de 2020, de THE CHILDREN'S MEDICAL CENTER CORPORATION: Una formulación farmacéutica para su uso en inducir pérdida de peso o reducir la grasa corporal, o una combinación de las mismas, en un paciente pre-obeso, obeso […]
Combinaciones y modos de administración de agentes terapéuticos y terapia combinada, del 8 de Abril de 2020, de ABRAXIS BIOSCIENCE, LLC: Una composición que comprende nanopartículas que comprenden paclitaxel y una albúmina, para uso en un método para el tratamiento de cáncer de pulmón en un individuo, en […]
Una composición para el tratamiento de la diabetes que comprende un análogo de oxintomodulina, del 11 de Marzo de 2020, de HANMI PHARM. CO., LTD.: Una composición para su uso en la prevención o el tratamiento de la diabetes, la composición comprende un conjugado de análogo de oxintomodulina como un ingrediente […]
Métodos para aumentar la producción de células madre hematopoyéticas y plaquetas, del 22 de Enero de 2020, de Janssen Pharmaceuticals, Inc: Una composición farmacéutica para su uso en un método de (i) proporcionar células madre hematopoyéticas a un sujeto; (ii) reducir el tiempo de injerto […]
Composición que comprende un péptido derivado de adinopectina, del 11 de Diciembre de 2019, de KNU-Industry Cooperation Foundation: Una composición farmacéutica adecuada para prevenir o tratar una enfermedad de la piel, que comprende un péptido seleccionado del grupo que consiste en los péptidos de las […]
POLIPÉPTIDOS, del 4 de Diciembre de 2019, de NOVO NORDISK A/S: Un polipéptido de acuerdo con la Fórmula (I): Xaa1-Cys-Xaa3-Thr-Ala-Thr-Cys-Ala-Thr-Gln-Arg-Leu-Ala-Xaa14-Phe-Leu- Xaa17-Xaa18-Ser-Ser- Xaa21- […]