HERPESVIRUS RECOMBINANTES DE PAVOS Y SUS USOS.
Un herpesvirus recombinante de pavos que comprende una secuencia de DNA extraño que en términos de la secuencia del DNA del virus de la la laringotraqueitis infecciosa (ILTV) consiste en una secuencia que codifica la glicoproteína I del virus de la laringotraqueitis infecciosa y la glicoproteína D del virus de la laringotraqueitis infecciosa dentro del fragmento EcoRI nº 9 del genoma viral del herpesvirus de pavos,
en donde la secuencia de DNA extraño que codifica dichas glicoproteínas es capaz de ser expresada en una célula hospedante infectada con el herpesvirus de pavos
Tipo: Patente Europea. Resumen de patente/invención. Número de Solicitud: E06026820.
Solicitante: SCHERING-PLOUGH LTD..
Nacionalidad solicitante: Suiza.
Dirección: Weystrasse 20, P.O.Box 6000 LUCERNE SUIZA.
Inventor/es: COCHRAN, MARK, D., JUNKER, DAVID, E., WILD, MARTHA, Singer,Philip.
Fecha de Publicación: .
Fecha Solicitud PCT: 9 de Agosto de 1995.
Clasificación PCT:
- C12N15/00 QUIMICA; METALURGIA. › C12 BIOQUIMICA; CERVEZA; BEBIDAS ALCOHOLICAS; VINO; VINAGRE; MICROBIOLOGIA; ENZIMOLOGIA; TECNICAS DE MUTACION O DE GENETICA. › C12N MICROORGANISMOS O ENZIMAS; COMPOSICIONES QUE LOS CONTIENEN; PROPAGACION, CULTIVO O CONSERVACION DE MICROORGANISMOS; TECNICAS DE MUTACION O DE INGENIERIA GENETICA; MEDIOS DE CULTIVO (medios para ensayos microbiológicos C12Q 1/00). › Técnicas de mutación o de ingeniería genética; ADN o ARN relacionado con la ingeniería genética, vectores, p. ej. plásmidos, o su aislamiento, su preparación o su purificación; Utilización de huéspedes para ello (mutantes o microorganismos modificados por ingeniería genética C12N 1/00, C12N 5/00, C12N 7/00; nuevas plantas en sí A01H; reproducción de plantas por técnicas de cultivo de tejidos A01H 4/00; nuevas razas animales en sí A01K 67/00; utilización de preparaciones medicinales que contienen material genético que es introducido en células del cuerpo humano para tratar enfermedades genéticas, terapia génica A61K 48/00; péptidos en general C07K).
- C12N15/09 C12N […] › C12N 15/00 Técnicas de mutación o de ingeniería genética; ADN o ARN relacionado con la ingeniería genética, vectores, p. ej. plásmidos, o su aislamiento, su preparación o su purificación; Utilización de huéspedes para ello (mutantes o microorganismos modificados por ingeniería genética C12N 1/00, C12N 5/00, C12N 7/00; nuevas plantas en sí A01H; reproducción de plantas por técnicas de cultivo de tejidos A01H 4/00; nuevas razas animales en sí A01K 67/00; utilización de preparaciones medicinales que contienen material genético que es introducido en células del cuerpo humano para tratar enfermedades genéticas, terapia génica A61K 48/00; péptidos en general C07K). › Tecnología del ADN recombinante.
- C12N15/12 C12N 15/00 […] › Genes que codifican proteínas animales.
- C12N15/19 C12N 15/00 […] › Interferones; Linfoquinas; Citoquinas.
- C12N15/24 C12N 15/00 […] › Interleuquinas.
- C12N15/26 C12N 15/00 […] › Interleuquina-2.
- C12N15/27 C12N 15/00 […] › Factores estimulantes de colonias.
- C12N5/10 C12N […] › C12N 5/00 Células no diferenciadas humanas, animales o vegetales, p. ej. líneas celulares; Tejidos; Su cultivo o conservación; Medios de cultivo para este fin (reproducción de plantas por técnicas de cultivo de tejidos A01H 4/00). › Células modificadas por introducción de material genético extraño, p. ej. células transformadas por virus.
- C12N5/20 C12N 5/00 […] › siendo uno de los integrantes de la fusión un linfocito B.
- C12N7/01 C12N […] › C12N 7/00 Virus, p. ej. bacteriófagos; Composiciones que los contienen; Su preparación o purificación (preparaciones de uso médico que contienen virus A61K 35/76; preparación de composiciones de uso médico que contienen antígenos o anticuerpos virales, p. ej. vacunas virales, A61K 39/00). › Virus, p. ej. Bacteriófagos, modificados por la introducción de material genético externo (vectores C12N 15/00).
Países PCT: Austria, Bélgica, Suiza, Alemania, Dinamarca, España, Francia, Reino Unido, Grecia, Italia, Liechtensein, Luxemburgo, Países Bajos, Suecia, Mónaco, Portugal, Irlanda.
PDF original: ES-2360298_T3.pdf


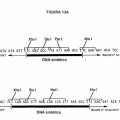
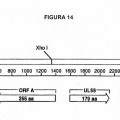
Fragmento de la descripción:
A lo largo de esta solicitud se mencionan varias publicaciones con números arábigos entre paréntesis. Las citas completas para estas publicaciones se encuentran al final de la descripción, inmediatamente antes de las reivindicaciones.
ANTECEDENTES DE LA INVENCIÓN
La capacidad de aislar DNA y clonar tal DNA aislado en plásmidos bacterianos ha expandido grandemente los métodos disponibles para preparar vacunas virales. Los métodos usados para conseguir la presente invención implican modificar las secuencias de DNA clonadas de varios agentes patógenos virales de animales, mediante inserciones, deleciones, cambios de bases únicos o múltiples e inserciones subsiguientes de estas secuencias modificadas en el genoma del virus. Una utilidad de la adición de una secuencia extraña se logra cuando la secuencia extraña codifica una proteína extraña que es expresada durante la infección viral del animal. El virus vivo resultante puede usarse luego en una vacuna para producir una respuesta inmunitaria en un animal hospedante y proporcionar protección al animal contra la enfermedad. Un virus con estas características se denomina un vector viral, porque se convierte en un vector vivo que llevará y expresará la proteína extraña en el animal hospedante. En efecto, llega a ser un sistema de administración elaborado para la(s) proteína(s) extraña(s).
Más específicamente, la presente invención se refiere al uso de herpesvirus de pavos (abreviadamente en lo sucesivo HVT por la expresión inglesa Herpesvirus of Turkey) como un vector viral para la vacunación de aves contra enfermedades. El grupo de herpesvirus comprende diversos agentes patógenos que infectan y causan enfermedades en cierto número de especies dianas: cerdos, ganado vacuno, pollos, caballos, perros, gatos etc. Cada herpesvirus es específico para su especie hospedante, pero todos ellos están relacionados en la estructura de sus genomas, su modo de replicación y en cierta extensión, en la patología que causan en el animal hospedante y en el mecanismo de la respuesta inmunitaria del hospedante a la infección por el virus.
La aplicación de las técnicas de DNA recombinante a los virus animales tiene una historia relativamente reciente. Los primeros virus que se manipularon por ingeniería genética han sido los que poseían los genomas más pequeños. En el caso de los papovavirus, debido a que estos virus son tan pequeños y no pueden acomodar mucho DNA extra, su uso en ingeniería genética ha sido como replicones defectuosos. La expresión del gen extraño de estos virus requiere un virus auxiliar de tipo natural y está limitada a sistemas de cultivos de células. Para los adenovirus, hay una pequeña cantidad de DNA no esencial que puede reemplazarse por secuencias extrañas. Los únicos DNA extraños que parece que se han expresado en adenovirus son los de los genes del antígeno T de papovavirus (Mansour et al., Proc. Natl. Acad. Sci. USA, 1985; Thummel et al., Cell, 1983; Scolnick, et al., Cell, 1981; Thummel et al., Cell 1981), y el gen de la timidina-quinasa del virus herpes simplex (abreviadamente en lo sucesivo HSV por la expresión inglesa Herpes Simplex Virus) (Haj-Ahmed y Graham, J. of Virology, 1986). Estas publicaciones no identifican las regiones no esenciales del HVT, en donde puede insertarse el DNA extraño, ni muestran cómo se logra la expresión de los genes extraños en el HVT, por ejemplo que secuencia de promotor y secuencia de terminación debe usarse.
Otro grupo de virus que se ha manipulado por ingeniería genética son los poxvirus. Un miembro de este grupo, el virus de la viruela de las vacas (denominado en lo sucesivo “virus vaccinia”), ha sido el objeto de mucha investigación sobre la expresión de genes extraños. Los poxvirus son virus que contiene DNA grandes que se replican en el citoplasma de la célula infectada. Dichos virus tienen una estructura que es única en el sentido de que no contienen ninguna cápsida que esté basada en simetría icosaédrica o simetría helicoidal. Los poxvirus son los que más probablemente han evolucionado de los microorganismos similares a bacterias por la pérdida de función y degeneración. En parte debido a esta singularidad, los avances hechos en ingeniería genética de los poxvirus no pueden extrapolarse directamente a otros sistemas virales, incluyendo los herpesvirus y el HVT. Las construcciones de virus recombinante del virus vaccinia se han hecho en un número de laboratorios que expresan los siguientes genes extraños insertados: gen de la timidina-quinasa del HSV (Mackett et al., Proc. Natl. Acad. Sci. USA, 1982; Panicali y Paoletti, Proc. Natl. Acad. Sci. USA, 1982, antígeno de superficie del virus de la hepatitis B (Paoletti et al., Proc. Natl. Acad. Sci. USA, 1984; Smith et al., Nature, 1983), gen de la glicoproteína D del HSV, gen de hemaglutinina del virus de la gripe (Panicali et al., Proc. Natl. Acad. Sci, USA, 1983; Smith et al., Proc. Natl. Acad. Sci. USA, 1983), gen del antígeno del virus de la malaria (Smith et al., Science, 1984) y gen de la glicoproteína G del virus de la estomatitis vesicular (Mackett et al., Science, 1986). Las características globales generales del DNA recombinante del virus vaccinia son similares a las técnicas usadas para todos los virus, especialmente en lo que se refieren a las técnicas de referencia (Maniatis et al., Molecular Cloning, 1982). Sin embargo en detalle, las técnicas del virus vaccinia no son aplicables al herpesvirus ni al HVT. La utilidad del virus vaccinia como vector de vacunas es un problema, debido a su relación cercana al virus de la viruela humana y su patogenicidad respecto a los seres humanos. Por tanto, el uso del herpesvirus HVT específico de hospedante es la mejor solución para la vacunación de las aves de corral.
Entre los herpesvirus de primate, solo el HSV de los seres humanos y, en una extensión limitada, el herpesvirus saimiri de los simios ha sido manipulado por ingeniería genética para contener secuencias de DNA extraño. El primer uso de DNA recombinante para manipular el HSV implicó clonar un trozo de DNA de la región de unión L-S en la región grande única de DNA del HSV, específicamente en el gen de timidina-quinasa (Moccarski et al., Cell, 1980). Este inserto no fue un trozo extraño de DNA, más bien fue un trozo que se presenta naturalmente de DNA de herpesvirus que se duplicó en otro lugar en el genoma. Este trozo de DNA no se manipuló genéticamente para expresar específicamente una proteína, y por tanto este trabajo no implica la expresión de proteínas en herpesvirus. La siguiente manipulación del HSV implicó la creación de deleciones en el genoma de virus mediante una combinación de técnicas de DNA recombinantes y de selección por timidina-quinasa. Usando este método se ha suprimido el gen alfa-22 del HSV (Post et al., Cell, 1981), y se ha suprimido una secuencia de 15.000 pares de bases de DNA de la región de repetición interna del HSV (Poffenberger et al., Proc. Natl. Acad. Sci. USA, 1981).
Los siguientes casos implican la inserción de genes que codifican proteínas en los herpesvirus: la inserción de la glicoproteína C del HSV en un mutante por deleción que ocurre naturalmente de este gen en HSV (Gibson y Spear,
J. of Virology, 1983); la inserción de la glicoproteína D del HSV de tipo 2 en HSV de tipo 1 (Lee et al., Proc. Natl. Acad. Sci. USA, 1982) sin manipulación de secuencias de promotor ya que el gen no es "extraño"; la inserción de antígeno de superficie del virus de la hepatitis B en el HSV bajo el control del promotor ICP4 del HSV (Shih et al., Proc. Natl. Acad. Sci. USA, 1984); y la inserción de la hormona de crecimiento bovina en el herpesvirus saimiri con un promotor de SV40 (el promotor no funcionó en este sistema y un promotor situado aguas arriba (es decir, la región que se extiende en dirección 5º) endógeno sirvió para transcribir el gen) (Desrosiers et al.,, 1984). Dos genes extraños adicionales (gen de ovalbúmina de pollo y el antígeno nuclear del virus de Epstein-Barr) se han insertado en el HSV (Arsenakis y Roizman, 1984) y la glicoproteína X del virus de la pseudo-rabia se han insertado en el HSV (Post et al., 1985).
Estos casos de deleción o de inserción de genes en herpesvirus demuestran que es posible manipular genéticamente genomas de herpesvirus mediante técnicas de DNA recombinantes. Los métodos que se han usado para insertar genes implican la recombinación homóloga entre el DNA viral clonado en plásmidos y el DNA viral purificado transfectado en la misma célula animal. Sin embargo, la extensión a la cual se puede generalizar la ubicación de la deleción y los sitios... [Seguir leyendo]
Reivindicaciones:
1. Un herpesvirus recombinante de pavos que comprende una secuencia de DNA extraño que en términos de la secuencia del DNA del virus de la la laringotraqueitis infecciosa (ILTV) consiste en una secuencia que codifica la glicoproteína I del virus de la laringotraqueitis infecciosa y la glicoproteína D del virus de la laringotraqueitis infecciosa dentro del fragmento EcoRI nº 9 del genoma viral del herpesvirus de pavos, en donde la secuencia de DNA extraño que codifica dichas glicoproteínas es capaz de ser expresada en una célula hospedante infectada con el herpesvirus de pavos.
2. Un vector para producir un herpesvirus recombinante de pavos insertando una secuencia de DNA extraño que en términos de la secuencia del DNA del virus la laringotraqueitis infecciosa (ILTV) consiste en una secuencia que codifica la glicoproteína I del virus de la laringotraqueitis infecciosa y la glicoproteína D del virus de la laringotraqueitis infecciosa en el genoma viral del herpesvirus de pavo, comprendiendo dicho vector una molécula de DNA bicatenario que consiste esencialmente en:
a) dicho DNA extraño en forma bicatenaria;
b) en un extremo del DNA extraño, DNA bicatenario de herpesvirus de pavos situado en un lado del fragmento EcoRI nº 9 de la región codificadora del genoma viral del herpesvirus de pavos; y;
c) en el otro extremo del DNA extraño, DNA bicatenario de herpesvirus de pavos situado en el otro lado del fragmento EcoRI nº9 de la región codificadora del genoma viral del herpes virus de pavos.
3. El vector de la reivindicación 2, en donde la secuencia de DNA extraño codifica un marcador detectable.
4. El vector de la reivindicación 3, en donde el marcador detectable es beta-galactosidasa de E. coli o beta glucuronidasa de E. coli.
5. Una vacuna útil para inmunizar un ave contra el virus de la laringotraqueitis infecciosa, que comprende una cantidad inmunizante eficaz del herpesvirus recombinante de pavos de la reivinidación 1 y un vehículo adecuado.
6. Una vacuna que comprende el herpesvirus recombinante de pavos de la reivindicación 1 para uso en inmunizar un ave contra el virus de la laringotraqueitis infecciosa.
7. Una célula hospedante infectada con el herpevirus recombinante de pavo de la reivindicación 1.
8. La célula hospedante de la reivindicación 7, en donde la célula hospedante es una célula aviar.
Patentes similares o relacionadas:
 CD52 soluble para su uso en el tratamiento o la prevención de la esclerosis múltiple o de la artritis reumatoide, del 29 de Julio de 2020, de THE WALTER AND ELIZA HALL INSTITUTE OF MEDICAL RESEARCH: Uno cualquiera o más de:
i) glucoproteína CD52 soluble, ii) una proteína de fusión que comprende la glucoproteína CD52 soluble como una primera proteína, y una segunda proteína;
[…]
CD52 soluble para su uso en el tratamiento o la prevención de la esclerosis múltiple o de la artritis reumatoide, del 29 de Julio de 2020, de THE WALTER AND ELIZA HALL INSTITUTE OF MEDICAL RESEARCH: Uno cualquiera o más de:
i) glucoproteína CD52 soluble, ii) una proteína de fusión que comprende la glucoproteína CD52 soluble como una primera proteína, y una segunda proteína;
[…]
Anticuerpo biespecífico o mezcla de anticuerpos con cadenas ligeras comunes, del 15 de Julio de 2020, de Jiangsu Alphamab Biopharmaceuticals Co., Ltd: Anticuerpo biespecífico o parte de unión a antígeno del mismo, en el que el anticuerpo biespecífico o la parte de unión a antígeno del mismo tiene una cadena […]
 Proteínas de dominio de fibronectina tipo III con solubilidad mejorada, del 24 de Junio de 2020, de BRISTOL-MYERS SQUIBB COMPANY: Un polipéptido que comprende restos de armazones a base de fibronectina que comprenden un 10º dominio de fibronectina tipo III (10Fn3) modificado, […]
Proteínas de dominio de fibronectina tipo III con solubilidad mejorada, del 24 de Junio de 2020, de BRISTOL-MYERS SQUIBB COMPANY: Un polipéptido que comprende restos de armazones a base de fibronectina que comprenden un 10º dominio de fibronectina tipo III (10Fn3) modificado, […]
PÉPTIDO DE MITICINA Y SU USO EN REGENERACIÓN CELULAR, del 4 de Junio de 2020, de CONSEJO SUPERIOR DE INVESTIGACIONES CIENTIFICAS: La presente invención se refiere a unos péptidos derivados de la micitina C y sus usos terapéuticos, más concretamente en la regeneración celular y/o […]
Péptido de miticina y su uso en regeneración celular, del 28 de Mayo de 2020, de CONSEJO SUPERIOR DE INVESTIGACIONES CIENTIFICAS: Péptido de miticina y su uso en regeneración celular. La presente invención se refiere a unos péptidos derivados de la micitina C y sus usos terapéuticos, más concretamente […]
Moléculas de unión de alta avidez que reconocen MAGE-A1, del 8 de Abril de 2020, de Max Delbrück Centrum für Molekulare Medizin (MDC) Berlin-Buch: Una construcción de reconocimiento de antígenos que es un receptor de células T (TCR), que comprende (i) una región variable de la cadena alfa […]
Moléculas de unión a ligando y usos de las mismas, del 25 de Diciembre de 2019, de Vegenics Pty Limited: Una molécula de unión a ligando que comprende un polipéptido de unión a ligando fusionado a un fragmento de dominio constante de inmunoglobulina, comprendiendo […]
Tratamiento de las enfermedades relacionadas con la apolipoproteína a1 por inhibición del transcrito antisentido natural a la apolipoproteína a1, del 17 de Octubre de 2019, de CuRNA, Inc: Un oligonucleótido que se dirige a un transcrito antisentido natural de apolipoproteína A1 para su uso como un compuesto terapéutico, donde el oligonucleótido […]